
Il rapporto con i patogeni
Se l’Africa è il luogo di origine e della prima evoluzione del genere Homo, per comprendere i rapporti coevolutivi tra Homo e patogeni è necessario approfondire l’argomento della distribuzione dei patogeni nel mondo, per capire se le malattie infettive siano distribuite a random o se esistano delle differenze caratterizzanti il continente africano.
Dato che l’indagine archeologica è impossibile per l’assenza di resti analizzabili, una conferma diretta sulla distribuzione dei patogeni nel periodo di interesse non è possibile, ma secondo Guegan e collaboratori (23) le inferenze dalle attuali distribuzioni permettono di dire che:
- la diversità delle specie patogene per l’uomo era ed è massima nelle zone tropicali e subtropicali.
- le specie di patogeni endemiche nelle zone temperate del mondo sono molto poche, mentre nelle zone tropicali sono presenti sia i patogeni endemici (patogeni, spesso zoonosi, con stadi esterni legati a vettori o a riserve, come gli elminti) sia quelli a distribuzione globale (di solito virus, batteri e funghi trasmessi direttamente, adattati alle popolazioni umane, con ciclo vitale interno all’uomo e quindi poco sensibili all’ambiente).
Ciò significa che le diverse popolazioni umane non sono state esposte allo stesso carico di malattie infettive, e che le popolazioni africane hanno avuto (e hanno) a che fare con una maggior diversità di patogeni.
I Cro Magnon erano probabilmente organizzati in piccoli gruppi egalitari di cacciatori e raccoglitori e, a differenza di H. neanderthalensis, avevano una dieta dominata dagli alimenti di origine vegetale. (24)
Come tutti gli ominidi, essi convivevano con parassiti con i quali si erano evoluti in Africa (dai macroparassiti come Enterobius, Ancylostoma, Uncinaria, Necator, ai microparassiti come Plasmodium responsabile della malaria e Flavivirus della febbre gialla) ed anche con parassiti di altri animali, ad esempio il Trypanosoma brucei rhodesiense della tripanosomiasi africana, il Leptospira della leptospirosi, la Brucella della brucellosi, la Salmonella della salmonellosi, lo Schistosoma della schistosomiasi, la Amoeba della dissenteria amebica, il Treponema pertenue, proveniente da animali o carne decomposta, che causa la framboesia, la Borrelia che porta la borreliosi, e la Yersinia pestis.
E’ probabile che, se non esenti da malattie, i primi Homo sapiens fossero comunque poco colpiti da malattie infettive, e soffrissero prevalentemente di ferite, traumi e di infezioni croniche a bassa intensità della pelle e del tratto gastrointestinale, le uniche che potevano mantenersi attive in popolazioni numericamente esigue, o perché duravano a lungo (dissenteria amebica) o perché potevano alternarsi tra ospiti diversi (schistosomiasi).(25)
Certamente non soffrivano di infezioni acute come morbillo o varicella, infezioni virali che o uccidono o rendono immuni e necessitano quindi di grandi numeri per mantenersi attive. Inoltre la maggior parte dei gruppi umani erano sempre in movimento, quindi non esistevano quelle riserve di focolai infettivi tipici degli insediamenti stabili che sono le latrine, la spazzatura e gli allevamenti.
Un caso di studio: la malaria
L’analisi delle frequenze di alcune malattie a base genetica ha dato indizi molto importanti proprio sul fondamentale ruolo selettivo/evolutivo delle malattie infettive. L’esempio più studiato è certamente quello del rapporto tra disordini dell’emoglobina e la malaria, che mostra come nonostante i fattori stocastici impliciti nella trasmissione della malaria, il rischio di infezione dipenda in buona parte da fattori predeterminati a livello genetico. (26)
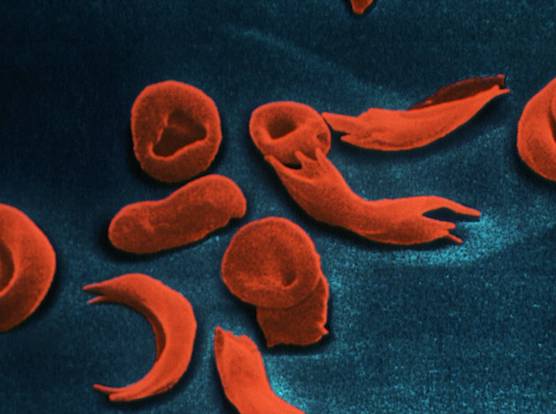
L’anemia falciforme risulta da una modificazione della subunità di tipo beta dell’emoglobina con formazione della emoglobina S (HbS) invece che la forma normale A (HbA). Negli omozigoti HbSS la HbS, quando viene ossidata, tende a precipitare e ad alterare la forma degli eritrociti, che divengono rigidi e distorti a falce (drepanociti), fragili, proni ad emolisi. I soggetti soffrono una elevata morbosità e mortalità, hanno aspettative di vita basse e raramente si riproducono.
L’allele modificato dovrebbe quindi essere estremamente raro o già scomparso, mentre si osservano frequenze molto elevate (più del 20%) nella fascia dell’Africa tropicale e frequenze meno elevate ma ancora superiori a quanto ci si aspetterebbe in Grecia, Turchia, India, Sicilia, ecc., mentre l’allele è assente in Nord America, Nord Europa, Australia.
Questa persistenza si potrebbe spiegare con una frequenza molto elevata di mutazione ricorrente, ma è più probabile che l’eterozigote HbAS abbia un vantaggio selettivo sugli individui “sani” HbAA. Questo vantaggio selettivo risulta evidente sovrapponendo le aree di persistenza dell’allele con quelle della distribuzione della malaria, aree che combaciano molto bene. Ed infatti si è scoperto che gli eterozigoti hanno ridotta prevalenza ed intensità della malaria rispetto agli omozigoti HbAA.

I parassiti della malaria (Plasmodium spp.) hanno più difficoltà a sopravvivere all’interno degli eritrociti anemici, probabilmente perché la loro azione pro-ossidante danneggia più facilmente l’eritrocita, causa una sua morte precoce e un rilascio di forme parassitarie immature che non sopravvivono all’esterno della cellula.
La stessa ipotesi di un vantaggio selettivo è stata avanzata anche per altre modificazioni patologiche dell’emoglobina, come alfa- e beta-talassemie, o per disfunzioni eritrocitarie, come ad esempio per il favismo, ovvero la deficienza dell’enzima Glucosio-6-fosfato deidrogenasi (G6PD). La deficienza di questo enzima chiave causa una reazione avversa a farmaci pro-ossidanti (l’emoglobina si ossida molto più facilmente, precipita e causa lisi dell’eritrocita) che si manifesta come una eccessiva distruzione di eritrociti. La ridotta capacità della cellula nel resistere allo stress ossidativo starebbe però alla base dell’effetto protettivo dalla mortalità da Plasmodium falciparum.
Come ha ben esposto Nina Etkin in un suo recente articolo la coscienza di questi legami evolutivi non è interessante solo dal punto di vista accademico, ma può funzionare come sapere applicato.(27) Comparare questi adattamenti biologici alla malaria agli adattamenti culturali, ad esempio la scelta delle piante medicinali o i comportamenti alimentari, ci può aiutare a spiegare perché tali adattamenti si siano presentati, e ci può aiutare a usare il dato etnobotanico come filtro per la ricerca di nuove piante utili.

L’autrice usa la pianta al momento più interessante per il trattamento della malaria, la Artemisia annua e la molecola artemisinina, mostrando come l’azione antimalarica derivi dal potenziale proossidante della molecola, che agisce sull’eritrocita e sul plasmodio, mimando in questo l’effetto di sensibilizzazione all’ossidazione delle anemie emolitiche.

L’autrice indica anche altri comportamenti come probabili adattamenti culturali di fronte alla malaria, come la tradizione est africana di fermentare la birra in recipienti ferrosi. La birra così ottenuta sarebbe carica di ferro, un fattore chiave nei processi ossidativi che faciliterebbe la lesione ossidativa agli eritrociti.
L’espansione
Con l’espandersi verso le nuove aree temperate, H. erectus e le altre specie di Homo si lasciarono indietro (in Africa) tutte le malattie con vettori o ospiti intermediari speciali e specifici del continente (tripanosoma, arborvirus, ecc.), mentre il clima più mite riduceva il carico di patogeni; se a queste differenze sommiamo il disgelo seguito all’ultima glaciazione (10.000 anni fa), si spiega forse la crescita demografica e la conseguente aumentata necessità di cibo che spinse verso la domesticazione degli animali e verso l’agricoltura. (28)
In questo quadro assume particolare rilevanza sanitaria il fatto che queste popolazioni assumessero sempre una grande varietà di cibi vegetali, ricchi di una grande diversità di nutrienti e di tossine vegetali, responsabili, come vedremo più avanti, della riduzione delle infezioni enteriche. (29)
Sempre questo quadro suggerisce che fosse ancora assente la figura dell’esperto guaritore, dell’esperto di piante medicinali e di riti, e che la gestione della salute ed il trattamento della malattia (vista ancora come un evento che si originava all’esterno del corpo, biologico e sociale) fosse collettivo e non segreto, folklorico e comunque comprendente un complesso di terapie razionali, sia chirurgiche sia erboristiche, usate per curare malattie semplici (diarrea, costipazione, ferite, ecc.) più un uso di tonici primaverili o altro che forse apportavano nutrienti. (30)
Come si vedrà più avanti, la “scoperta” dell’agricoltura, con la possibilità di discriminare tra piante spontanee e piante coltivate, piante alimentari e piante medicinali, permette la individuazione di soggetti esperti e di conoscenze segrete, limitate agli esperti, esoteriche.
La conquista del mondo
I movimenti migratori che hanno portato H. sapiens a conquistare il mondo sono conosciuti nelle loro linee più generali.
Nell’arco temporale del “grande balzo in avanti”, dopo la conquista dell’Eurasia meridionale, H. sapiens arriva in Australia e Nuova Guinea (unite al tempo a causa della glaciazione) tra i 30.000 e i 40.000 anni fa (iniziando l’estinzione della megafauna australasiana), con quello che è stato probabilmente il primo utilizzo di imbarcazioni per superare grandi distanze (intorno agli 80 km). Circa 20.000 anni fa l’uomo conquista le terre fredde della Siberia, probabilmente contribuendo all’estinzione del Mammut e del rinoceronte lanoso. E’ probabile che solo le maggiori capacità di H. sapiens rispetto ad H. erectus e H. neanderthalensis abbiano permesso questo passaggio.
L’ultima grande massa continentale ad essere conquistata è stata l’America. Approfittando di favorevoli condizioni climatiche, è probabile che intorno a 12-000 anni fa i primi coloni siano arrivati in Alaska, e che nel giro di mille anni queste popolazioni siano arrivate in Patagonia. La Groenlandia dovrà aspettare il 2000 a.C. (31)
Se la parte principale della dieta di Homo sapiens arcaico era costituita dai vegetali (lo indicherebbero le strie dei denti comparabili a quelle dei vegetariani contemporanei, i cestini per la raccolta di vegetali nel tardo Paleolitico, i fitoliti indicanti uso di cereali, il rapporto Stronzio/Calcio delle ossa che si innalza nel Mesolitico), con il passare del tempo egli diviene sempre più attivo nel procacciarsi la carne, passando da scavenger passivo a scavenger attivo e cacciatore, e gli strumenti, specie quelli utilizzati per la macellazione delle carcasse animali, si fanno più sofisticati a mano a mano che cresce la competizione con i grossi carnivori.
Certamente l’utilizzo più massiccio della carne come alimento energetico facilita l’apporto di principi nutritivi atti a sostenere l’encefalizzazione e quindi l’ominazione.
A questo periodo risalgono altri importanti ritrovamenti di indizi sull’uso delle piante da parte dell’uomo. I resti trovati nei siti Neolitici degli abitanti dei laghi dell’Europa centrale indicano coltivazione o raccolta di ca. 200 specie diverse di piante (ad es. papavero da oppio, Papaverum somniferum L. — Papaveraceae).

Il maggior consumo di cibi ad elevata densità e d’origine animale ha probabilmente migliorato lo status nutrizionale di Homo sapiens ma ha anche cambiato il suo rapporto con foglie e composti allelopatici, ed è probabile che questi cambiamenti abbiano avuto un effetto sull’equilibrio tra status nutritivo, organismi patogeni e proprietà positive e negative dei composti attivi. Il cambiamento di dieta, infatti, potrebbe aver reso da un lato meno necessario l’utilizzo di foglie (energeticamente povere) e dall’altro aver reso possibile un loro consumo più elevato in caso di necessità (perché un organismo ben nutrito detossifica più facilmente gli xenobiotici, ovvero i composti chimici farmacologicamente attivi esogeni introdotti con la dieta).
Forse è qui, con lo sganciamento parziale dell’uomo dalla necessità di ingerire piante tossiche, e con l’inizio del lungo processo che avrebbe portato alla domesticazione di alcune piante, che si ha per la prima volta la possibilità di parlare di medicina e non solo di comportamenti di automedicazione. Perché il disaccoppiamento della frazione nutritiva da quella tossica permette di individuare due soggetti fino a questo momento fortemente sovrapposti: le piante alimentari e le piante medicinali, ed è possibile ingerire, coscientemente, composti allelopatici a scopo curativo.
——————————
Note
23. Guegan J-F, Prugnolle F, Thomas F (2008) “Global spatial patterns of infectuous diseases and human evolution”. In S.C. Stearns & J.C. Koella (eds.) Evolution in Health and Disease. Second Edition. Oxford University Press
24. Kiple K.F. “The ecology of disease”. in W.F. Bynum e R. Porter 1993 op. cit. pp. 357-381. Anche se la presenza di asce e coltelli di pietra e di segni da taglio sui denti indicano un utilizzo di carne, le strie sui denti e la loro qualità estremamente simile a quelle dei vegetariani odierni indica una dieta prevalentemente vegetariana (Consiglio e Siani 2003 op. cit. )
25. In mancanza di dati archeologici, la fonte più importante di inferenze sul passato sono le condizioni di vita odierne delle ultime popolazioni di cacciatori raccoglitori; essi sono ben nutriti rispetto ai vicini coltivatori, e di solito in salute (Vickers W.T. “The health significance of wild plants for the Siona and Secoya”. In Etkin, N.L. (Ed.), 1994 op. cit. pp. 143-165), ed i loro problemi parassitari ed infettivi sono probabilmente in equilibrio con la popolazione (Kiple 1993 op. cit. ).
26. Ma, come hanno mostrato Mackinnon MJ, Mwangi TW, Snow RW, Marsh K, Williams TN (2005) “Heritability of malaria in Africa”. PLoS Med 2(12): e340, i fattori genetici dell’ospite sembrano contare per il 25-33% della variabilità totale nella suscettibilità, e solo una piccola percentuale di questa variazione sembra legata ai geni più conosciuti e studiati, rafforzando l’ipotesi che la suscettibilità alla malaria sia sotto il controllo di molti geni differenti, e di fattori non genetici sempre predeterminati, che si articolano in maniera complessa con i fattori genetici.
27. Etkin, N (2003) “The co-evolution of people, plants, and parasites: biological and cultural adaptations to malaria”. Proceedings of the Nutrition Society, 62:311-317
28. Diamond 1997 op. cit.
29. Johns 1990 op. cit.; Vickers 1994 op. cit. ; Kiple 1993 op. cit.
30. Anche in questo caso ci si rifa ai agli studi effettuati sulle ultime popolazioni di cacciatori raccoglitori, che utilizzano rimedi per molti problemi: ferite, fratture, slogature, dolore, problemi di pelle, febbre, raffreddore, tosse, diarrea, mal di testa, ecc. Le piante venivano e vengono consumate come infusi, forse ancora prima come pianta fresca o secca ingerita tal quale.
31. Diamond 1997 op. cit.











