Nel post precedente (qui) ho iniziato a parlare del problema della valutazione e dell’utilizzo dei dati etnobotanici e storici. Ho analizzato uno dei punti critici del processo di indagine, cioè la valutazione quantitativa della segregazione delle piante medicinali nelle famiglie botaniche, concludendo che questa segregazione esiste e sembra indicare una scelta non casuale delle piante da parte dei gruppi umani. Quali poi siano le ragioni per questa scelta è naturalmente un problema più complesso, e che quasi certamente non permette risposte mono-causali.
E’ stato proposto [1] che gli schemi di classificazione che osserviamo nelle e tra le società siano il risultato di:
- predisposizioni cognitive universali risultanti dall’evoluzione.
- l’oggettiva struttura tassonomica del bioma locale che non possiamo fare a meno di riconoscere.
- interpretazioni culturali relative.
- meri artefatti dei metodi di ricerca.
Il sapere tradizionale
Ma prima di affrontare questo argomento è necessario evocare il convitato di pietra di questa discussione, cioè il “sapere tradizionale” (o sapere ecologico tradizionale, o sapere locale, come è stato variamente definito). Fino ad ora, e nel post precedente, per comodità di trattazione ho usato questo termine (d’ora in poi ST) come se non esistessero ambiguità rispetto alla sua definizione, alla sua estensione, alle sue origini. Ma è utile soffermarci di più sulla sua articolazione all’interno della ricerca in etnobotanica, perché è essenziale per l’esplorazione del rapporto tra dati storico-etnobotanici ed efficacia biologica, ed inoltre perché è proprio sull’interpretazione del termine e sul sull’utilizzo da fare del “sapere tradizionale” che spesso casca l’asino della discussione erboristica/fitoterapica.
Partiamo intanto con una definizione preliminare tratta da un articolo di Victoria Reyes-García del 2010. L’autrice definisce il ST o i sistemi di sapere tradizionale come: “il sapere sulle risorse e le dinamiche dell’ecosistema, e le pratiche di gestione ad esso associate esistenti tra i membri della comunità che, giornalmente e per lunghi periodi di tempo, interagiscono per il proprio benessere e per la coesistenza con l’ecosistema stesso”. Secondo l’autrice il ST include non solo informazioni sugli usi umani di piante ed animali, ma anche sistemi di classificazione, osservazioni sull’ambiente locale e un sistema di utilizzo e gestione delle risorse, oltre a credenze su esseri non umani, sovrannaturali, e sulle relazioni che essi intrattengono con la società. In definitiva il sapere tradizionale deve essere inteso come in modo di comprendere il mondo o la “cultura”.Esso rappresenterebbe un tratto culturale adattivo per le popolazioni che favorisce la loro sopravvivenza, la produzione di cibo, la costruzione di ripari e in genere il controllo sulle proprie vite. Verrebbe sviluppato e continuamente adattato ad un ambiente in continuo cambiamento, e verrebbe passato di generazione in generazione non isolato bensì strettamente intrecciato a valori culturali ed etici.
L’altro
Vista questa definizione, torniamo a bomba al problema dello status dei dati storici ed etnobotanici. Quello che a mio parere è uno dei problemi più diffusi in questo ambito è la tendenza a polarizzare i termini del dibattito seguendo una antica tendenza della discussione sull’altro o sull’alterità, cristallizzata dall’antropologia settecentesca intorno all’alternativa “buon selvaggio” (preservato dalla corruzione della civiltà grazie alle sue doti naturali) o “cattivo selvaggio” (immerso nella miseria e depravazione a causa della sua ignoranza e pigrizia).
Succede infatti spesso che l’altro erboristico (sia esso il medico antico che seguiva i dettami della sua tradizione di medicina colta, o sia esso il guaritore contemporaneo che in una società distante utilizza rimedi della foresta) venga costruito secondo una immagine idilliaca, con un discorso impressionistico e poco rigoroso. Si richiama, con modalità talvolta ingenua e nostalgica, un passato nel quale si potrebbero ritrovare le origini più vere, la vicinanza alla fonte, qualcosa di “originario” che il presente, il progresso e la modernità ci hanno tolto. Oppure si cerca nelle popolazioni esotiche e nelle loro culture “la silhouette mobile e vaga di un selvaggio che, più vicino di noi alla natura, avrebbe rifiutato in anticipo tutto ciò che ci opprime e del quale si troverebbe ancora traccia, ricordo o testimonianza nella foresta amazzonica o nel deserto australiano” (vedi ad esempio Marc Augè).
Termini che vengono spesso usati in questo contesto sono tradizione, radici, origine, memoria, ecc., ma raramente gli autori che li usano si fermano a pensare al perché essi vengano usati, ed in che modo. Questo ragionamento schematico e dicotomico, etnocentrico al rovescio, contribuisce ad imprigionare l’altro, a sottometterlo alla nostra rappresentazione. Spesso in questo dibattito si fa uso del termine e della metafora delle radici [2], che richiama il tema delle origini, che in quanto origini sembrano più importanti, imprescindibili, contenenti in nuce ciò che da esse si svolge [3]. Ma come dice Galimberti:
“Essere più vicini alla fonte non significa custodire qualcosa di “originario”, ma essere semplicemente all’inizio di un processo: la storia, che si compie facendosi, e non abolendola dissetandosi alla fonte. All’inizio c’è solo l’avvio, e non il senso nascosto, o il silenzio custodito da ciò che in seguito si dispiegherà. E solo il rifiuto del mondo che viviamo può far ritenere che il mondo antico, con il suo corredo di simboli, disponga di segni più veri. Ma rifiuto e nostalgia sono i moti dell’anima di chi disabita il mondo che per sorte si trova ad abitare, non sono certo criteri di giudizio, ne tantomeno sentieri di verità”.
Quantificazione
Fatta questa doverosa premessa circa l’atteggiamento metodologico da tenere quando siamo confrontati da dati storici ed etnobotanici, rimane l’aspetto tecnico, interno diciamo all’etnobotanica. Cosa significa il termine sapere tradizionale o i suoi cognati? E’ possibile quantificarlo? E se si, come?
La quantificazione del sapere tradizionale è stata tentata attraverso l’utilizzo di indici di misura dell’importanza culturale delle piante, i cosiddetti indici di Importanza Culturale Relativa [RCI]. Questi indici quantitativi si usano in etnobotanica per comparare usi e importanza culturale dei differenti taxa. Sono stati usati in vari studi e la loro utilità risiede in prima istanza nel fatto che su di essi si possono effettuare analisi statistiche di vario tipo per comparare diverse specie tra loro, zone vegetazionali, habitus, taxa, ecc. Essi inoltre permettono di ottenere valori numerici integrabili ad altri indici (tassonomici, fitochimici, ecc.) utilizzabili per comparazione trans-culturali e per testare differenti ipotesi.
Gli indici RCI si sono moltiplicati negli ultimi anni, ma mi soffermo in questo momento su tre gruppi di indici che sono particolarmente importanti.
1. Indici di totalizzazione degli usi
Semplici indici enumerativi, che elencano usi e non usi per ogni pianta. Un esempio è UT

Dove UT è pari alla semplice sommatoria di tutti gli usi conosciuti per ogni specie, che possono essere categorizzati per utilità, per taxon vegetale o per tipo vegetazionale. Questi metodi non tentano di misurare i livelli di importanza per i differenti usi e non tengono conto del consenso tra gli informatori. Il taxon più importante è semplicemente quello con le maggiori citazioni d’uso. Dal punto di vista della possibilità di catturare la complessità dell’oggetto sapere tradizionale (e della misura dell’importanza culturale) questi indici non offrono molto, e dal punto di vista della rilevanza statistica e della possibilità di testare delle ipotesi sono i meno efficaci. Non registrando la provenienza dell’informazione (da quale informatore) non permettono di valutare la variabilità intraculturale. Non tenendo in conto l’esistenza di livelli diversi di importanza, il metodo è eccessivamente sensibile all’intensità della campionatura (una pianta con molti utilizzi tutti poco importanti nel gruppo studiato peserebbe di più di una pianta con meno utilizzi ma molto più importanti). Per ovviare almeno in parte a questi deficit, sono stati sviluppati dei metodi più sofisticati, come quelli detti di assegnazione soggettiva dell’importanza.
2. Indici di assegnazione soggettiva
Indici simili a quelli precedenti, ma con modificatori di importanza basati sulle valutazioni e sulla conoscenza del contesto da parte del ricercatore. Questi indici pesati permettono la misura dei gradi di importanza ma introducono un pregiudizio del ricercatore che rende più difficile l’utilizzo degli indici, ad esempio nella analisi bibliografica. Un esempio è quello del Valore d’Uso (UVs)
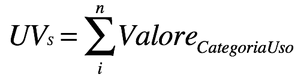
UVs deriva da UT ma somma i punteggi generati dal ricercatore per ogni utilizzo (importante = 1 punto; poco importante = 0,5 punti).
Gli indici che sono stati più influenti nei decenni recenti sono però senza dubbio stati gli indici di consenso.
3. Indici di consenso tra gli informatori
Indici basati sul consenso tra gli informatori, con pesi dati agli usi generati dal ricercatore o dall’informatore. Questi indici si basano sulla teoria del consenso culturale (CCT), una teoria antropologica sviluppata per stimare la risposta culturalmente corretta in differenti domini del sapere tradizionale. La CCT parte dagli assunti che esista una risposta culturalmente corretta per ogni domanda [4], che il sapere consista nell’accordo tra informatori [5], e che la probabilità che un informatore risponda in maniera corretta ad una determinata domanda sia il risultato della sua competenza in quel dominio di sapere [6]. Si tratta quindi di un metodo che misura il sapere in termini di congruenza con un modello esplicativo proprio del gruppo studiato. Da ciò deriva l’ipotesi che maggiore la salienza di una pianta o di un suo utilizzo in una comunità, maggiore sarà la frequenza di citazione della pianta.
Qui sotto riporto alcune delle formule più usate.
1. Valore d’uso delle specie per un informatore:
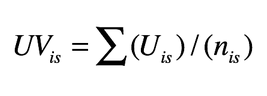
Dove UVis = numero di usi menzionati per la specie s dall’informatore i; n is = numero di interviste nelle quali l’informatore i menziona un uso per la specie s.
2. Valore d’uso di una specie per tutti gli informatori
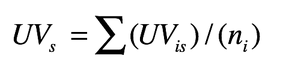
Dove ni = numero totale di informatori intervistati per la specie s. Questo indice misura il numero di usi attribuiti ad un taxon relativo al numero di informatori che citano il taxon, cioè misura l’importanza del taxon se per importanza intendiamo la sintesi degli usi. Alcuni autori hanno infatti riscontrato una correlazione positiva tra numero di informatori che considerano importante una specie s, l’indice UV, e il numero di utilizzi, ed hanno concluso che l’importanza è una sintesi della molteplicità degli usi di un taxon. Altri autori preferiscono legare l’importanza sia al numero di usi sia a quanto bene è conosciuta la pianta.
UVs può essere ritenuta rilevante quando tanti informatori citano molti utilizzi, ma può essere fuorviante se vi è un solo informatore che cita molti utilizzi, oppure se una pianta è oggettivamente molto importante, ad esempio una pianta simbolo stesso della società studiata, ma ha pochi usi. Teoricamente gli indici basati sul consenso degli informatori dovrebbero favorire quei taxon con un elevato livello di consenso in una cultura, ma un indice come UV è ancora molto crudo.
3. Metodo del punteggio dell’informatore
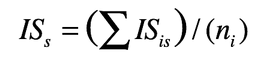
All’uso di una pianta viene assegnato dall’informatore un punteggio variabile da 0,5 (utilizzabile ma sub-ottimale) a 1,5 (quasi ottimale) in cinque categorie. Questi punteggi vengono sommati per ottenere un punteggio finale, per una singola intervista, nella forchetta 0-7,5. ISis si calcola come il punteggio medio per tutte le interviste, e ISs si ottiene come la media tra i valori d’uso per tutti gli informatori.
Criticità
Tutte queste tecniche condividono l’assunto che gli indici misurino il sapere tradizionale, e che l’estensione del sapere tradizionale su una pianta in una data cultura ci dia una misura indiretta dell’importanza culturale percepita di tale pianta in maniera più oggettiva degli altri indici. Inoltre, quando gli indici vengono utilizzati a scopo euristico, per delimitare taxa specifici per la ricerca etnofarmacologica, ci si basa sull’assunto che vi sia un legame tra importanza culturale, sapere tradizionale, efficacia percepita e potenziale farmacologico.
Ognuno di questi passaggi presenta delle criticità.
- Il rapporto tra indici RCI ed efficacia percepita potrebbe non essere lineare. Vi è ad esempio il ruolo della disponibilità delle piante che può confondere questo rapporto.
- Esistono molte discrepanze tra dati ottenuti con metodi differenti, discrepanze che rivelano come il sapere tradizionale comprenda molte dimensioni e campi differenti, rendendo difficile incapsularlo in un’unica definizione derivante da un unico metodo.
- Esistono discrepanze tra “sapere attivo” (quali piante vengono usate dalle persone) e “sapere passivo” (cosa le persone sanno sull’utilizzo delle piante)
- La correlazione tra sapere e consenso potrebbe non essere lineare e semplice, e potrebbe dipendere:
4.1. dal fatto che stiamo studiando piante medicinali o piante alimentari o piante medicinali e alimentari.
4.2. Da chi decidiamo di intervistare come informatore (guaritore, bambini, uomini, donne, ecc.). Gli etnofarmacologi si sono quasi sempre concentrati sui guaritori professionisti perché li ritenevano i più profondi conoscitori della materia, ma se professionisti e gente comune non condividono lo stesso corpus di sapere, allora restringere la ricerca ad un solo gruppo limita e rende pregiudiziale il tipo di informazione raccolta
4.3. Dalla differenza tra sapere condiviso e sapere idiosincratico
Casagrande mette anche in dubbio l’assunto che le piante usate più di frequente siano anche quelle percepite come più efficaci. L’autore arriva a questa conclusione dopo avere riscontrato nel suo studio sul campo che:
- L’abbondanza di una specie è collegata alla frequenza d’uso, mentre la frequenza d’uso non è collegata all’efficacia percepita; secondo l’autore questo indica che le prime piante ad essere utilizzate in caso di malattia sono le più disponibili, le più abbondanti nella vicinità delle zone abitate, a prescindere dalla loro efficacia. Le piante più efficaci (efficacia emica) vengono usate solo in caso le prime non risolvano il problema. Visto che la severità dei sintomi può scemare anche senza intervento, è probabile che in molti casi le piante più efficaci non verranno utilizzate.
- L’efficacia percepita e, in misura minore, la frequenza d’uso, sono correlati alla distribuzione del sapere, ma l’organizzazione sociale, i fattori cognitivi individuali e distribuiti, i processi di trasmissione culturale strutturata e random danno forma al processo di disseminazione. A parte poche piante e malattie molto comuni, la distribuzione del sapere sarà spesso random o non prevedibile.
Questi risultati mettono in crisi il postulato di molta ricerca quantitativa che suppone che frequenza d’uso, distribuzione del sapere e/o consenso tra informatori si correli bene con l’efficacia percepita, e mette quindi in dubbio la correlazione tra indici RCI ed efficacia emica. I risultati metterebbero anche in dubbio l’ipotesi che le piante più disponibili siano più utilizzate perché conterrebbero più composti interessanti (come voleva Moerman): le piante più disponibili verrebbero più usate semplicemente perché sono più disponibili.
Sarebbe quindi meglio misurare direttamente l’efficacia percepita, ma anche il legane tra efficacia percepita e potenziale farmacologico non è netta, perché le informazioni variano tra le popolazioni, è quindi necessario comparare diverse comunità in aree geografiche diverse e preferibilmente lontane.
Quindi, secondo Casagrande i rapporti tra frequenza d’uso e abbondanza di una pianta non sono indicatori forti dell’efficacia percepita, mentre il consenso lo è, e comunque l’efficacia percepita ha un legame debole con il poitenziale famacologico se non viene analizzata a livello transculturale (vedi il prossimo post).
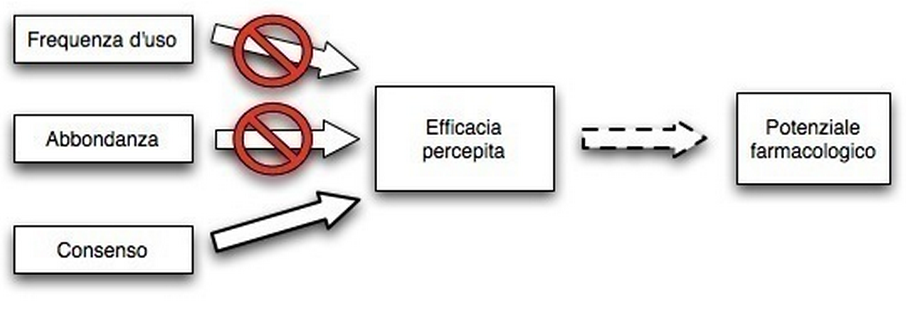
Questo non significa che, secondo Casagrande, le piante usate dalle popolazioni non siano mai efficaci, ma che gli schemi di distribuzione del sapere non rappresentano una corrispondenza ottimale tra bisogni indotti dalla malattia e i composti fitochimici disponibili nell’ambiente.
Il prossimo post si concentrerà su due articoli che descrivono due applicazioni molto diverse tra loro degli indici, ma che ci possono servire per capirne meglio l’utilità.
————————————————————————————————————————————————————————————————–
Note
[1] Casagrande, David 2004 Ethnobiology lives! Theory, Collaboration, and Possibilities for the Study of Folk Biologies. Reviews in Anthropology, vol. 33, pp. 351 to 370
[2] Una metafora molto potente perché mentre nessuno ha mai visto una tradizione o una identità, tutti hanno visto delle radici.
[3] Lo stesso concetto di tradizione viene presentato come monolitico e fisso. In effetti se non fosse presentato in questo modo non potrebbe svolgere il compito che gli viene richiesto, quello di “ancorare” le pratiche presenti ad un passato “naturale” che le giustifichi. Ma questa visione, di nuovo piuttosto ingenua, non rende conto del carattere costruttivo e dinamico della tradizione e della memoria collettiva che hanno bisogno di una serie di cornici di riferimento a carattere sociale che ne condizionano fortemente i contenuti. Al mutare dei quadri sociali muta la memoria. La memoria collettiva e la tradizione culturale si creano attraverso un processo di ricostruzione artificiale (vedi il Palio di Siena, la topografia leggendaria dei Vangeli in Terra Santa, la costruzione delle differenze etniche tra Hutu e Tutsi) (Halbwachs (1987) La memoria collettiva, Unicopli, Milano). Proprio gli antichi sembravano ben avvertiti del carattere dinamico e non monolitico della tradizione, incapace da sola a fondare una identità: Cicerone (De legibus, 2, 16, 40) racconta che un’ambasciata ateniese si recò a Delfi per chiedere ad Apollo quali riti sacri mantenere e quali no. L’oracolo rispose: eos quae essent in more maiorum, cioè “quelli conformi al costume degli antenati”, ovvero “tenetevi alla vostra tradizione”. Ma gli ateniesi tornarono poco dopo per ottenere maggiori delucidazioni sul significato dell’indicazione: infatti il costume degli antenati era mutato molte volte, quale era la tradizione che dovevano ritenere quella valida? Al che Apollo rispose: “la migliore”, rivelando che la tradizione non è ne fissa ne monolitica, cambia e si costruisce, e alla fine la scelta si basa su criteri utilitaristici, non sull’antichità.
[4] Nel senso che la realtà culturale, definita come la risposta data dalla maggior parte delle persone, è la stessa per tutti gli informatori (o le fonti di informazione, in caso si lavori su testi.
[5] Il livello di accordo tra informatori riflette il loro accordo congiunto
[6] Se consideriamo la competenza come la percentuale di riposte corrette.